The human retina
The retina (at the back of the eye) is composed of several types of cells, as well as blood vessels.
Surprisingly, light has to pass through at least 2 layers of neurones before reaching the
photoreceptor cells: 'rods' and 'cones'.
These are so called because of the shape of the outer segment. Here is a stack of membranes, containing a pigment which makes them light-sensitive.
Rod cells are responsible for monochrome ("black and white") vision, whilst cones give colour vision. They also vary in sensitivity, as explained below.
Impulses from the photoreceptors are passed via synapses to intermediate
bipolar neurones - so called because each has two processes on either side of the cell body - and these pass impulses on to
ganglion cells which feed into the optic nerve. The left and right optic nerves pass into the optic chiasma, effectively crossing to the other side of the brain and into the thalamus, which passes impulses on to the visual cortex, at the back of the cerebral hemispheres of the brain, where the nervous information is interpreted as vision.
Rods and cones are typically connected differently to their bipolar cells.
One bipolar cell normally receives impulses from several rod cells, and sends impulses on to more than one ganglion cell.
On the other hand, each cone cell normally sends its impulses on to a single bipolar cell and then on to a single ganglion cell.
This means that rod cells are particularly useful in low light levels, but they do not give such clear and high-definition vision as cones, which require higher levels of illumination.
The distribution of cones and the pattern of nerves in the fovea are responsible for vision with the highest definition -
visual acuity - as well as discriminating colours of light reflected from objects under view.
It also explains why it is necessary to (turn the head and) line up the visual axis of the eye with the object being studied, in class or reading a book, for example. However it is known that in the dark it is easier to see stars more clearly by looking slightly to one side.
Photochemistry
The pigment in rods is called rhodopsin, and its molecule consists of a protein part - opsin - attached to retinal - a derivative of vitamin A. When this absorbs light energy, its molecule changes shape (from 11-cis to all-trans) and this alters membrane permeability, producing a generator poential.
The cone cells themselves are not differently coloured - they just respond to different colours of light
Similarly, cone cells have retinal attached to different opsin proteins.
There are actually three types of cone cell. Each one has a slightly different opsin, so that they respond to different wavelengths of light, corresponding to different colours of light: blue, green and red. This is called the
trichromatic theory of colour vision.
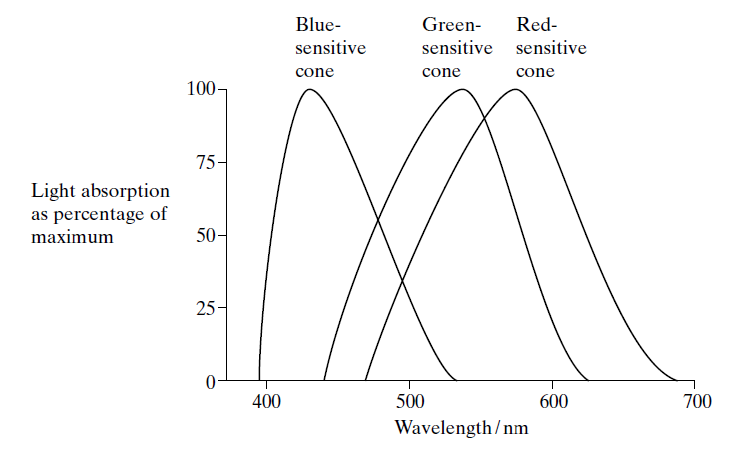
This graph shows the percentage of light of different wavelengths absorbed by the three
types of cone cell found in the retina of the human eye. Light of different colours is detected by stimulation of different combinations of these cells. This has a parallel to the system by which computer users can specify colours of pixels and text on screen by reference to numbers for rgb (
red, green and blue components of light produced).
Layers of cells in the retina
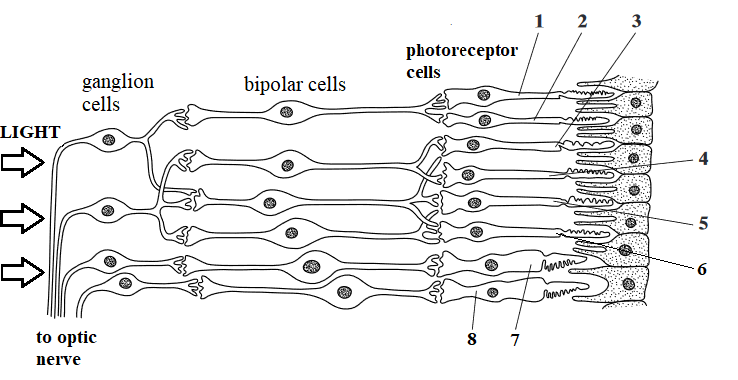 Section through the retina, showing types of neurones
Section through the retina, showing types of neurones
Photoreceptors labelled 1-6 are rods, 7 & 8 are cones
Rod cells 1 and 2 share the same ganglion cell and neurone, so light falling on both of these only registers as a single spot.
Rod cell 3 has a separate neurone so when light falls on 2 and 3, two separate spots may be seen.
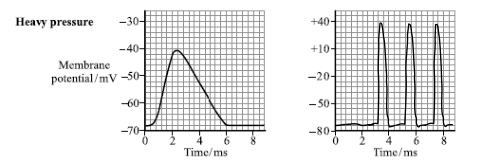
When a small amount of light falls on a cell, e.g. 3 , it may not be seen because the receptor potential is sub-threshold.
However, if a similar amount of light falls on several, e.g. rod cells 3, 4, and 5, this may be seen, because collectively they are above threshold, i.e. they cause sufficient depolarisation to stimulate an action potential in the next cell. This is an example of spatial summation, with 3 receptors releasing enough neurotransmitter to generate an impulse.
Don't use the term "messages" or "signals" when referring to nerve impulses
This phenomenon is called
retinal convergence, as light falling on several nearby rod cells results in each of them producing impulses which are
brought together to a single bipolar cell and outgoing neurone, resulting in a single action potential leaving the eye in one 'nerve fibre' of the optic nerve, which is interpreted in the brain as a single point of light.
Large numbers of
cone cells similar to 7 and 8 are found in the fovea. These give higher visual acuity than rod cells 1-6, because each photoreceptor connects to a single bipolar cell and ganglion, and pass action potentials out along separate nerve fibres, so they are each are registered in the brain as a distinct point of light.
They also give colour vision, because there are 3 different types of cones, each responding to different colours/wavelengths of light
All in the back of the eye
The retina consists of about 120 million rods, widely spread out, and about 5 million cones. About 2000 cones (and practically no rods) are concentrated in an area in the centre of the optical axis at the back of the eye. This region of maximal sensitivity is called the
fovea centralis , and it is served by about half the nerve fibres that make up the optic nerve (consequently feeding on to half the visual cortex). There is less obstruction by blood vessels and overlying connective neurones in the central region.
There are several zones, varying in the packing density of cones and bipolar cells. The term
macula may be used interchangeably with fovea, or it may represent a slightly larger area. Macular degeneration is an (age related) condition in which cellular changes in this area lead to reduced clarity of (central) vision.
There are no photoreceptors where the optic nerve leaves the eye, seen below as the optic disc. This is the 'blind spot'.
Can you see through this?
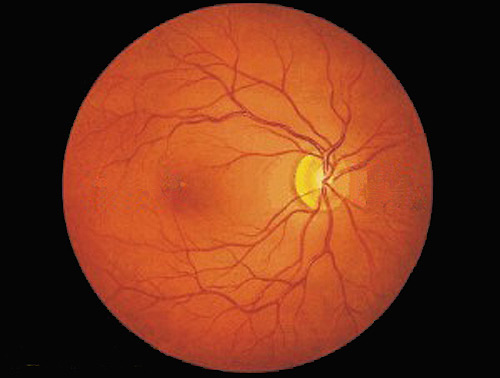 A photograph of the back of the eye ('fundus') showing the surface of the retina, and the dark orange colour is mainly due to visual pigments.
A photograph of the back of the eye ('fundus') showing the surface of the retina, and the dark orange colour is mainly due to visual pigments.
The fovea is the small blob in the indistinct dark area on the left.
The optic disc (composed of axons from ganglia) shows as a light circular zone on the right, with blood vessels emerging.
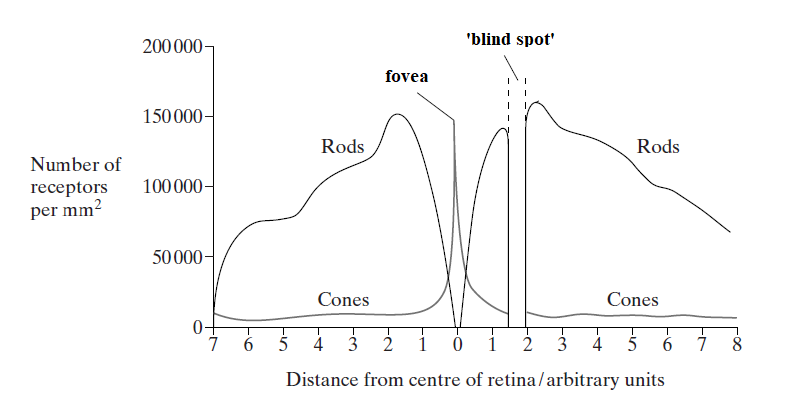
The distribution of rod cells and cone cells across the retina of a human eye
N.b. Even though there are high numbers of rods in some regions of the retina, they do not give such good resolution as cones do, because of the way their associated neurones are 'connected'.
Colour blindness
There are several forms of colour blindness, differing in the range of colour loss and severity.
Red-green colour blindness is a fairly common sex-linked condition due to a recessive allele which causes problems in distinguishing between hues in the red-green area.
The most common form
deuteranomaly results from a change in the green-sensitive pigment and is only a minor inconvenience to some. The frequency in males is about 6-8%.
A more severe form of red-green colour blindness
deuteranopia results from the complete absence of one set of cones - the green-sensitive ones.
This is also known as
Daltonism - after
John Dalton - who introduced the atomic theory in Chemistry. The frequency in males is about 1%.
When analysing allele and phenotype frequencies by the Hardy-Weinberg methodology, it must be modified to reflect the limitation caused by the X chromosome:
in males allele frequencies for affected and unaffected are identical to phenotype frequencies, typically p and q, not squared,
whereas in females phenotype frequencies are the 'normal' 3 categories: p2, 2pq and q2, as there are 2 copies of the X chromosome.
Total colour blindness
achromatopsia is caused by the absence of all cone cells, resulting in monochromatic vision, in which everything is seen in shades of grey - like on a black and white television. This is inherited as an autosomal recessive condition, and is very uncommon - in the USA the frequency of individuals affected is about 0.003%.
However on the small
tropical Micronesian island of Pingelap, the frequency is said to be about 10%, a consequence of a founder effect/genetic bottleneck resulting from the repopulation of the island following a tornado in 1775 which greatly reduced the population. All affected individuals today can trace their ancestry back to
one individual who survived.
It is said that the eyes of some affected individuals are sensitive to daylight sun levels, but they are at an advantage when night fishing.
There are some inconsistencies in online accounts, such as the suggestion that 30% of the island's population (of 250) are carriers.
Hardy-Weinberg analysis suggests q2 = 0.1 so q = 0.32, and p = 0.68. The 2pq value for heterozygote frequency should therefore be (2 x 0.32 x 0.68) = 0.4352 - in other words 43.5%.
But perhaps there is another explanation, such as immigrants upsetting the equilibrium.
What do you see?
 The answer to the Ultimate Question of Life, the Universe, and Everything?
The answer to the Ultimate Question of Life, the Universe, and Everything?
Colour blindness is a sex-linked condition, so it is more common in males.
If you can see two numbers, you have normal colour vision.
If you see only one number clearly, you probably have '
green colour blindness' or '
red colour blindness'.
If you see one number clearly and another faintly, you probably have a mild version '
mild green colour blindness' or '
mild red colour blindness'.
For more tests see web references below.

 Home
Home