The light-independent reaction
Do not call this the dark reaction, or fall into the trap of assuming that it occurs later on at night.
It follows directly after the light-dependent reaction.
It is helpful to keep track of the number of carbon atoms (and phosphate groups) in the various compounds. The chemical conversions are all part of a cycle - usually called the
Calvin cycle.
Carbon dioxide reacts with
ribulose bisphosphate (RuBP)
to form two molecules of
glycerate 3-phosphate (GP). This reaction is catalysed by the enzyme RuBisCo.
 Glycerate 3-phosphate
Glycerate 3-phosphate is
reduced to
triose phosphate (3-C sugar with 1 phosphate), using ATP and reduced NADP from the light-dependent reaction.
Most of the triose phosphate (10 out of 12 molecules) is used in a multi-stage process to
regenerate RuBP, so as to repeat the process of accepting carbon dioxide.
The rest of the triose phosphate is the product of the light-independent reaction, and 2 molecules of it are produced for every 6 molecules of CO
2 which enter the cycle.
This is converted to
useful organic substances required by the plant, e.g.
glucose (a 6-C sugar) which may be converted (by polymerisation) into other carbohydrates, e.g. cellulose for cell walls or stored as starch.
It can also be converted into other classes of organic compound, e.g. lipids, amino acids and then proteins and nucleic acids.
The Calvin cycle
Melvin Calvin (1911�1997) was an American biochemist at the University of California, Berkeley.
Along with Andrew Benson and James Bassham, Calvin discovered the plant metabolic cycle which bears his name, sometimes together with the others.
He (alone) was awarded the 1961 Nobel Prize in Chemistry.

Calvin used this 'lollipop apparatus' in which he grew the green alga
Chlorella in a liquid containing carbon dioxide labelled with radioactive
14C and he took extracts after illuminating it for different (short) periods of time.
Two-dimensional chromatograms were prepared and these were covered with photographic paper and left (in the dark) for some time before being processed in the normal way to develop a photographic image - an autoradiogram. Dark splodges showed the position of radioactive organic compounds resulting from the reaction, which were then identified. Reducing the time (and increasing the intensity of light) allowed the sequence of products to be determined.

It is interesting to note that in addition to carbohydrate products, amino acids have been formed in the later sample.
What is meant by a two-dimensional chromatogram?
>
Chromatogram that is run in one direction, then dried, rotated through 90°, and run again, probably with a different solvent
What are its advantages?
>
To separate overlapping compounds
The LIR in more detail
The following events take place within the
stroma - a larger fluid-filled space outside the thylakoids which is surrounded by the two outer chloroplast membranes - each of which is itself a lipid bilayer, with embedded proteins. The lipid compositions of these membranes differ from one another and from most other cell membranes which are mostly phospholipid. In fact the outer membrane is about 48% phospholipids, 46% galactolipids and 6% sulpholipids, whereas the composition of the inner membrane is 16% phospholipids, 79% galactolipids and 5% sulpholipids. This is quite similar to composition of the thylakoid membrane.
The chemical reactions which take place in the stroma are a cycle of enzyme-controlled covalent conversions and do not rely directly on
electron transfer which necessitates a membrane-bound chain of redox carriers.
Carbon dioxide fixation
 Ribulose 1,5 bisphosphate (RuBP)
Ribulose 1,5 bisphosphate (RuBP) is a double phosphate ester of the ketopentose sugar ribulose. This is similar to ribose - a 5-carbon sugar - but it has a >C=O keto group at carbon 2, and it is 'linear' not ring-shaped as it has two phosphate groups, one on each end of the molecule.
RuBP is carboxylated (it reacts with carbon dioxide) to form an unstable compound, 3-keto-2-carboxyarabinitol 1,5-bisphosphate.
The enzyme RuBisCO (Ribulose Bisphosphate CarbOxylase) which catalyses this reaction is said to be the most common protein in the world as it makes up 20-25% of the mass of protein in most leaves and it is produced at a rate of about 1000 kg/second on earth.


The diagrams above show molecules in zigzag skeletal format,
but the Carbon coming in from CO2 is shown as a C

The unstable 6-carbon, 2 phosphate compound splits in the middle to form 2 molecules of
glycerate-3 phosphate - GP - a 3-carbon, 1 phosphate compound similar to glycerol but with a carboxylic acid group - shown on the right as -COO
- (conjugate base of glyceric acid).
Reduction

GP is further
phosphorylated using ATP to form
glycerate 1,3 bisphosphate (G1,3BP) (1,3 phosphoglycerate).
The next stage is effectively the same as in glycolysis (steps 6&7), but in reverse.
G1,3BP is reduced by reacting with reduced NADP which provides protons and electrons.

The resulting products are
glyceraldehyde 3-phosphate (G3P, also abbreviated to PGA, PGAL or GALP and others!) and inorganic phosphate.
G3P is possibly best known as
triose phosphate (TP), and some of this is converted into useful compounds for the plant.
Typically 2 x G3P out of 12 molecules in the cycle can be converted into
fructose 1,6 bisphosphate, fructose 6 phosphate, then glucose 6 phosphate, then glucose.
Regeneration of RuBp

The rest of the triose phosphate (10 out of 12 molecules) undergoes molecular rearrangement - in 8 stages - to form
ribulose monophosphate - ribulose 5-phosphate, Ru5P. This is then phosphorylated to form
ribulose 1,5 bisphosphate again, so the cycle can continue.
Both RuBP and ADP have two phosphate groups.
Why do we write RuBP as ribulose bisphosphate, but ADP as adenosine diphosphate?
>
In ADP both phosphates are attached (in a row) to the same part of the molecule, in RuBP there are are 2 different parts - at opposite ends of the molecule
Photorespiration - the downside of RuBisCO
As well as catalysing the (carboxylation) reaction between RuBP and carbon dioxide, the enzyme RuBisCO can also catalyse the (oxygenation) reaction between RuBP and oxygen.
Presumably the molecules of CO
2 and O
2 are similar in size and shape so that both can fit into the active site of RuBisCO.
This reaction, known as photorespiration, is wasteful as it results in the production of 2-phosphoglycolate at well as glycerate 3-phosphate (2 molecules of which are normally produced). The 2-phosphoglycolate is processed via a different cycle before being recycled into RuBP to re-enter the Calvin cycle, and carbon dioxide is lost in the process.
This is a drawback to the normal ('
C3') photosynthesis, as described above. It is more marked in conditions of low carbon dioxide concentration - especially when stomata are closed due to drought - or higher temperature.
Some plants use alternative methods of trapping carbon dioxide, using the enzyme
phosphoenolpyruvate carboxylase, which produces
4-carbon compounds that are exported to another part of the leaf with a low oxygen concentration where CO
2 is released to be accepted by RuBisCO in the normal way, avoiding oxygenation.
Some of the world's most economically important (tropical, grasslike) crops - sweetcorn or maize (
Zea mays), sugarcane (
Saccharum officinarum), sorghum (
Sorghum bicolor), and millets (several species) - use this
C4 photosynthesis.
Most succulent plants and cacti use another alternative: Crassulacean Acid Metabolism
CAM in which CO
2 is captured at night when it is cooler and there is less water loss via open stomata, and it is converted into C4 compounds and malic acid which is stored in the vacuole.
During daytime they release CO
2 from their reserves to perform photosynthesis when stomata are closed and water conservation mechanisms operate.
Evolution of photosynthesis
Early conditions on planet Earth were very different than today:
Many environments were anaerobic and reducing, rather like hydrothermal vents today.
There was practically no oxygen in the atmosphere, but a higher amount of carbon dioxide
This meant that there was no ozone layer and much more ultraviolet light.
Carbon dioxide dissolved to produce hydrogencarbonate ions.
Originally there must have been bacteria with an established respiration process involving movement of electrons between a number of redox molecules like haem and cytochromes. Within one of these a single photosystem evolved, as magnesium replaced iron in the porphyin ring of haem and the pigment chlorophyll was formed, and this absorbed energy from light.
This early photosystem probably took electrons from hydrogen sulphide or iron, and using chlorophyll passed them into the respiratory cycles enabling organic compounds like sugars to be produced from CO
2.
Another version of the photosystem emerged which probably took excited electrons from chlorophyll and used them to move protons across the membrane and create ATP before returning the electrons to chlorophyll. This is similar to cyclic photophosphorylation. Both of these systems would differ somewhat and evolved to be more efficient in different environments.
It is thought that (possibly following gene exchange) some bacteria had both systems, but they had a mechanism which allowed them to use only one at a time, avoiding unnecessary synthesis of intermediates: the first provided organic compounds for growth when hydrogen sulphide was available, and the second gave ATP for maintenance when it was unavailable. This is called the "redox switch hypothesis".
Nowadays, several groups of bacteria perform photosynthesis but most have only photosystem I or photosystem II; only cyanobacteria have both systems available, and in some species they switch off PS II when performing nitrogen fixation, which is inhibited by oxygen.
Conceivably depletion of raw materials meant that these bacteria were subject to selection pressure.
Manganese, which is quite common on the ocean floor today, absorbs ultraviolet radiation which could be destructive to organic molecules within cells, so it would act as a sort of antioxidant screen to this radiation. In doing so it emits electrons which would be absorbed by chlorophyll which has lost electrons as it absorbed light energy.
It is thought that cyanobacteria used manganese to split water, which is a much more widely available chemical resource.
Furthermore it is thought that bypassing the redox switch enabled the two systems to work in synchrony.
The resulting continuous electron flow could then be passed from one photosystem to the other, and used to produce both ATP and reduced NADP.
This double photosystem with the manganese-based oxygen-evolving complex became the standard combination when chloroplasts were formed by the coalescence of two cells, forming an early single-celled organism like an alga, from which many-celled plants presumably evolved. Cyanobacteria and higher plants have transformed Earth's atmosphere by adding oxygen obtained by splitting water, and this has allowed a number of aerobically respiring organisms to evolve.
See 'endosymbiont theory' (link below).
Most of the previous section dealt with the light-dependent reaction.
Several versions of the light-independent reaction (C
3, C
4, CAM) have been mentioned above, but earlier versions probably used respiration pathways (running in the opposite direction to 'normal').
For example, green sulphur bacteria are currently often found in deep waters, near to hydrothermal vents which emit a dull light as well as various mineral ions which provide reducing conditions resulting in an anoxic environment. Unsurprisingly, they are anaerobic.
They employ PS I for photosynthesis, using several bacteriochlorophylls which absorb light at 720-750 nm but the P840 chlorophylls at the reaction centre emit electrons which are passed to ferredoxin for reduction of NADP. Electrons are repaid following the oxidation of sulphide, resulting in the accumulation of elemental sulphur. The fixation of carbon dioxide is achieved by a reaction which is the complete reverse of the tricarboxylic acid (Krebs, citric acid) cycle, so they use a respiratory pathway.

 The oak tree has a canopy of photosynthetic tissue, and this can be a food source for a variety of moth caterpillars, which are eaten by birds like bluetits, which are preyed upon by other birds like sparrowhawks.
The oak tree has a canopy of photosynthetic tissue, and this can be a food source for a variety of moth caterpillars, which are eaten by birds like bluetits, which are preyed upon by other birds like sparrowhawks.
 The oak tree leaf is an organ of photosynthesis. The veins bring in water and the leaf's flattened shape exposes it efficiently to sunlight. It also means that carbon dioxide has only to diffuse a short distance from below to reach the cells inside the leaf.
The oak tree leaf is an organ of photosynthesis. The veins bring in water and the leaf's flattened shape exposes it efficiently to sunlight. It also means that carbon dioxide has only to diffuse a short distance from below to reach the cells inside the leaf.
 The mesophyll - the main photosynthetic tissue - is in the middle of the leaf between the upper and lower epidermis.
The green dots are chloroplasts.
The mesophyll - the main photosynthetic tissue - is in the middle of the leaf between the upper and lower epidermis.
The green dots are chloroplasts.

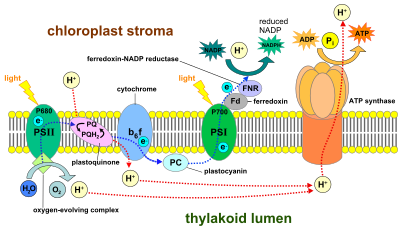
 This diagram shows the location of proteins within the thylakoid membrane and their relationship to the lumen and the stroma
This diagram shows the location of proteins within the thylakoid membrane and their relationship to the lumen and the stroma
 Chlorophyll has a square porphyrin section, very similar to the haem section within haemoglobin, but it contains a magnesium ion whereas haem contains an iron ion.
Chlorophyll has a square porphyrin section, very similar to the haem section within haemoglobin, but it contains a magnesium ion whereas haem contains an iron ion.















 Home
Home