|
Site author Richard Steane
|
The BioTopics website gives access to interactive resource material, developed to support the
learning and teaching of Biology at a variety of levels.
|
|
|
|
Digestion and absorption
The digestion process
Digestion is the process of hydrolysing complex biological compounds into simpler ones by breaking bonds between sections of the molecules.
It is effectively the conversion of polymers into monomers. Often polymers are digested into dimers, which are then converted into monomers in a separate stage.
Reminder: hydrolysis involves the addition of -H and -OH from water, and it is achieved by the use of specific enzymes, collectively known as hydrolases.
Large biological molecules are hydrolysed to smaller subunits that can cross cell membranes and be absorbed from the digestive system.
Once they have entered cells they can be built up into other compounds (assimilated) or more fully broken down to release energy. It is important to get the processes of digestion and respiration into perspective. In particular digestion does not of itself release energy - it gives cells the opportunities to process foods in preparation for respiration.
The essence of digestion in the mammalian alimentary canal is that complex insoluble substances are converted into simpler soluble compounds, which can be moved in solution by the circulatory system.
This means that glucose can be delivered to respiring cells alongside oxygen. As such, this topic is a natural partner to 'Mass transport in animals'. (Links below)
Within the digestive system, molecules are first acted on by enzymes secreted into the mass of food after it has been chewed and moistened by digestive juices. These enzymes are described as extracellular or luminal, i.e. acting in the centre of the mass of food in the gut, away from the cells that produce them.
The first enzymes secreted do not always fully complete the hydrolysis, leaving smaller sections of the food molecules to be acted on by other enzymes.
Digestion is often completed by 'membrane-bound' enzymes that are attached to the apical surfaces of cells lining the alimentary canal.
Final products of digestion can cross membranes by facilitated diffusion using specific carrier proteins and they can then be absorbed into associated blood vessels.
Digestion of carbohydrates
Polysaccharides (carbohydrate polymers)
Starch is the main complex carbohydrate in diet, but at this level it is probably best considered as the compounds amylose and amylopectin.
Glycogen is a similar compound, of animal origin.
These are all built from a large number of glucose units, usually bonded by α-1→4 glycosidic linkages, but in amylopectin and glycogen occasional α-1→6 bonds permit branching so there are plenty of 'loose ends' accessible to amylase enzymes.
Enzymes involved
Amylase is secreted by the salivary glands and the pancreas and it mixes with food that has been ingested.
It breaks α-1→4 glycosidic bonds that are 2 units from the exposed ends, but not next to a branch point.
It therefore produces predominantly maltose - a dimer of glucose, as well as maltotriose - a trimer of glucose - and α-limit dextrins- 5-6 glucose residues with a branch point.
Oligosaccharides (mostly carbohydrate dimers)
The disaccharide maltose is the main product of the first stage of polysaccharide digestion.
This is then further hydrolysed by maltase.
Two other disaccharides are sucrose - basic sugar from cane or beet - and lactose - 'milk sugar'.
Sucrose is glucose joined to fructose by an α1→ β2 linkage
Lactose is galactose joined to glucose with a β1→4 linkage
Enzymes involved - disaccharidases
The enzyme maltase is attached to the brush borders of cells lining the villi in the ileum. This hydrolyses the single α-1→4 glycosidic linkage in maltose, releasing 2 glucose units.
Isomaltase is another membrane-bound enzyme that breaks α-1→6 bonds between glucose residues.
Other enzymes hydrolysing specific disaccharides are sucrase and lactase.
Starch is about 25% amylose and 75% amylopectin, although there is some variation between different species and varieties of food plants.
Amylose is a 'straight-chain' α-1,4 linked polymer of hundreds or thousands of glucose units, but it forms into a helical tube-like structure. A 100-residue section would be about 30nm in length. Amylose is less easily digestible than amylopectin, on account of its regular fairly tight coiling, and it is only exposed for hydrolysis at the end (ends?) of the strand.
Amylopectin is a branched chain polymer with similarly shaped (but shorter) α-1,4 linked glucose sections radiating out every 20-30 glucose units in 3 dimensions from a large number of branch points, each of which has a a-1,6 linkage. Each molecule contains in total up to 200,000 (or more?) units of glucose.
Glycogen from animals is similar to amylopectin in structure but it is smaller , as it is more highly branched (8-12 glucose units between branches). It may consist of up to 60,000 glucose units. It is found in liver and muscle, so it is a carbohydrate of interest to carnivores!
Enzymes in pairs
These disaccharidase enzymes tend to be produced as dimers, or at least they have 2 catalytic domains - separate active sites. Their polypeptide chains have been sequenced and their structures have been investigated. Molecular explanations are often based on the location of the ends of the chain: N-terminal refers to the end of the polypeptide chain with a free -NH2 (amino) group, and the C-terminal end has a free -COOH (carboxylic acid) group.
What has happened to amino groups and carboxylic acid groups between the two ends?
> They have been used to form peptide bonds ( -CONH-) in the polypeptide chain.
Maltase-glucoamylase consists of two subunits, one - the N-terminal catalytic domain - targetting maltose, and the other - C-terminal domain - acting against a broader spectrum of glucose oligomers.
These are anchored to the brush-border on the outer membrane of cells lining the small intestine, by means of an O-glycosylated stalk - glucose attached to serine in the N-terminal domain.
Digestion of proteins
Polypeptides
Proteins can be considered to be linear chains of amino acids.
Since there are 20 amino acids, the number of combinations of amino acids in proteins is endless. There is only one type of bond involved - the peptide bond - but different sidegroups on amino acids can effectively shield the links, making them more resistant to enzyme action, or conversely they may act as access points for enzymes to attach and carry out hydrolysis.
Enzymes involved
Different proteolytic enzymes (proteases, peptidases) break the polypeptide chains at different positions:
Endopeptidases break peptide bonds of nonterminal amino acids (i.e. within the molecule), usually after specific amino acids. As a result, they produce several shorter polypeptide chains, but not single amino acids. They do produce many more 'free ends' for exopeptidases (See below) to act on.
In some cases endopeptidases are very specific in their action, but others may act on a variety of target amino acid sequences. Often the target amino acids can be summed up by simple references to sidechain chemistry, e.g. acidic groups, basic groups, or aromatic groups. Proline (an imino acid, not a true amino acid) sometimes assists, and sometimes prevents hydrolysis of the chain.
These enzymes also show quite marked pH 'preferences', and these generally match with their location within the alimentary canal.
Examples:
Pepsin
Trypsin
Chymotrypsin
All of these are secreted as inactive forms or precursors ('zymogens'): pepsinogen, trypsinogen, chymotrypsinogen. This prevents damage to the organs that secrete them, or to the wall of the alimentary canal (which is protected to a certain extent by a layer of mucus).
Pepsinogen is activated by hydrochloric acid and pepsin already in the stomach.
Trypsinogen is produced and stored in the pancreas, and after it is secreted into the duodenum as part of pancreatic juice it is activated by the enzyme enteropeptidase (enterokinase), which is found in the intestinal mucosa. This splits off a 15 amino acid sequence and the remaining section forms the active form trypsin, which can also activate more trypsinogen by removing the same segment.
Chymotrypsinogen is activated by trypsin.
Exopeptidases cleave a bond at one end of the (shortened) polypeptide chain, releasing a single amino acid or dipeptide.
Carboxypeptidases act at the C-terminal end (breaking one peptide bond just behind the terminal amino acid with a carboxylic acid group -COOH.
They are present in pancreatic juice. However they will not hydrolyse a dipeptide.
Aminopeptidases act at the N-terminal end (breaking one peptide bond just behind the terminal amino acid with an amino group -NH2.
They may be found in the brush border of cells lining the ileum.
Dipeptides
Although exopeptidases produce single amino acids, they sometimes do not hydrolyse dipeptides. A number of fairly undefined dipeptides will remain.
Enzymes involved
Membrane-bound dipeptidases in the ileum cleave dipeptides into their two component amino acids prior to absorption.
Individual amino acids can cross the cell membrane into lining cells as there are carrier proteins for amino acids whereas dipeptides cannot cross the cell membrane as there are no carrier proteins for them.
More detail about endopeptidases
| Enzyme | Target residues |
| Pepsin | cuts before Leu, Phe, Trp or Tyr, unless preceded by Pro |
| Trypsin | cuts after Arg or Lys, unless followed by Pro |
| Chymotrypsin | cuts after Phe, Trp, or Tyr, unless followed by Pro |
*Glutamyl endopeptidase I
- "V8 protease" | cuts after Glu or Asp |
*isolated from
Staphylococcus aureus strain V8
A polypeptide chain under attack
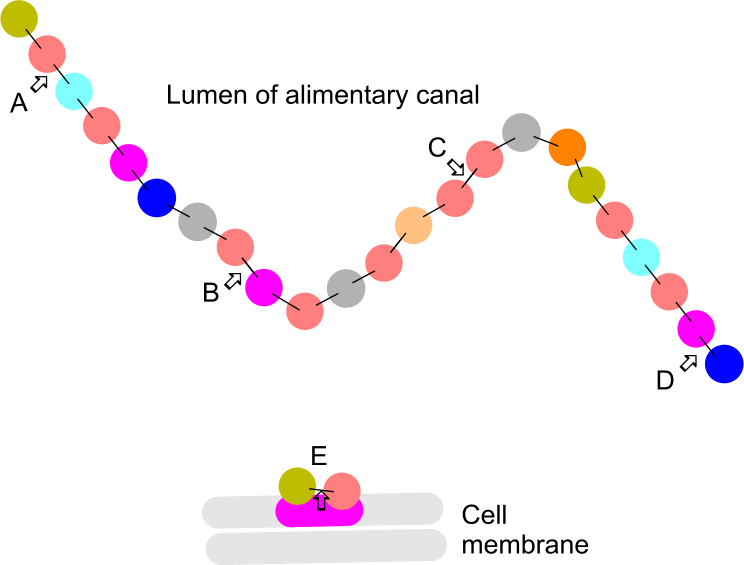 What (type of enzyme) is acting in the following positions?
What (type of enzyme) is acting in the following positions?
A >
exopeptidase
B >
endopeptidase
C >
endpeptidase
D >
exopeptidase
E >
dipeptidase
Digestion of lipids
Triglycerides (fats and oils) differ from carbohydrates and proteins in that they are not built up from a potentially large number of sub-units, so they are not polymers.
The action of lipase is assisted by bile salts (principal components of bile, produced in the liver, stored in the gall bladder) which emulsify fats, breaking them down into small droplets and increasing the surface area for contact with the water-soluble enzyme.
Digestion of a lipid molecule usually results in the release of one molecule of monoglyceride (glycerol attached to one fatty acid) and two fatty acids, or one glycerol molecule and three fatty acids.
[Fatty acids are not broken down in the digestion process so their hydrocarbon chains are not reduced in length at this stage.]
Enzymes involved
Pancreatic lipase, secreted in the pancreatic juice, is the main enzyme that performs this function.
Gastric lipase also hydrolyses fats and oils in the acidic conditions of the stomach.
Colipase is a co-enzyme required for optimal activity of pancreatic lipase. It is secreted by the pancreas in an inactive form, procolipase, which is activated in the intestinal lumen by trypsin. Its function is to prevent the inhibitory effect of bile salts on the hydrolysis of dietary long-chain triglycerides.
Absorption of the products of digestion
Structure of an intestinal villus
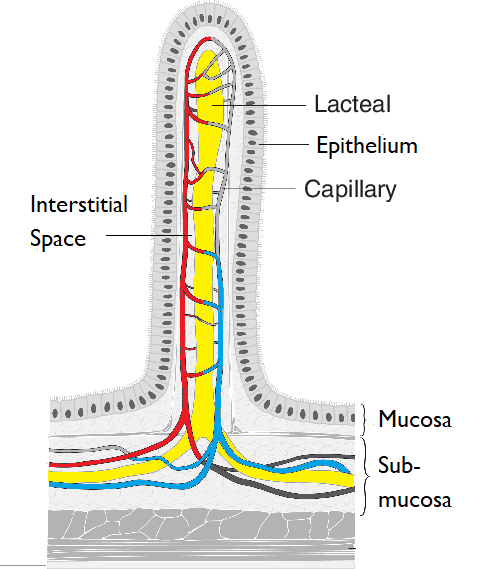 Modified from:
DeSesso, John & Jacobson, Catherine & Williams, Amy. (2012). Anatomical and Physiological Parameters that Influence Gastrointestinal Absorption. 10.1002/9780470921920.edm022.
Modified from:
DeSesso, John & Jacobson, Catherine & Williams, Amy. (2012). Anatomical and Physiological Parameters that Influence Gastrointestinal Absorption. 10.1002/9780470921920.edm022.
The ileum of mammals is lined with thousands of finger-like projections called
villi, which increase the surface area for absorption of digestion products. These are covered with an epithelium of cells called enterocytes which themselves have
microvilli forming a
brush border on the side projecting out into the lumen of the alimentary canal. Each villus has in its centre a network of blood capillaries and a single lymphatic capillary (sometimes known as a
lacteal).
Carbohydrate digestion products and protein digestion products are water soluble, and there are mechanisms to collect and absorb these into the bloodstream.
.
Blood carrying aqueous digestion products (glucose and amino-acids) is taken via the
hepatic portal vein to the liver.
Here carbohydrate may be converted into glycogen for storage (glycogenesis), respired for energy, or released into the systemic circulation.
Amino acids may be released into the bloodstream, and any excess can be deaminated in the liver, to give urea and the rest of the molecule can be respired to provide energy.
Lipid digestion products, although also water soluble, tend to reassociate into molecules which need to be processed inside a cell of the intestinal epithelium before they are taken away by a separate system: the lymphatic system. Eventually this deposits encapsulated lipid into the blood system.
The human digestive system
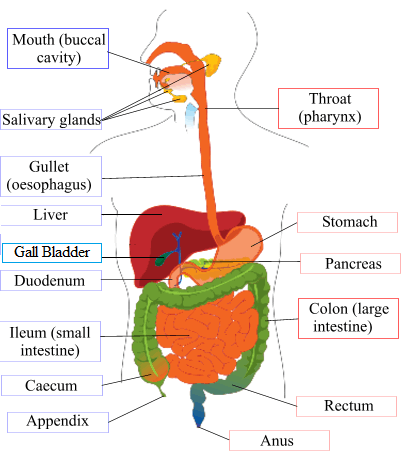 I thought I would put in this diagram to set this topic in the perspective of the human body
I thought I would put in this diagram to set this topic in the perspective of the human body
Absorption of amino acids and of monosaccharides
Processes involved in absorbing amino acids and monosaccharides into a capillary from the
small intestine
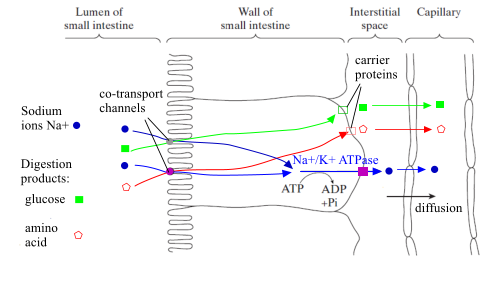 What name is given to the part of the lining cell shown by a zigzag line at the left of the diagram?
What name is given to the part of the lining cell shown by a zigzag line at the left of the diagram?
>
brush border/microvilli
What advantage does this provide in the absorption process?
>
gives larger surface area/more channel proteins
Amino acids and monosaccharides are absorbed by secondary active transport, which has several stages.
Sodium ions enter cells lining the ileum together with either of these products of digestion, via a channel protein, of which there are several sorts.
This process - facilitated diffusion - is an example of
co-transport.
Sodium ions are pumped out of the cell and into the interstitial fluid using an ATP-powered protein pump. This establishes a concentration gradient of Na
+ ions between the lumen and the lining cell. Arguably it could be said that it also establishes a concentration gradient of digestion products.
Each product of digestion leaves the cell via a different carrier protein/permease channel.
They then enter the blood capillary by diffusion. This is assisted by the steady movement of blood which maintains a concentration gradient by taking away the digestion products.
Micelles and chylomicrons in the absorption of lipids
Stages in the digestion and absorption of lipids in the ileum
Not to scale
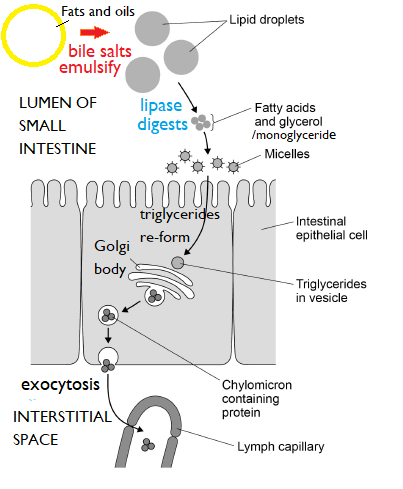
Monoglycerides, glycerol and fatty acids are all more water-soluble than triglycerides, although they still have a hydrophobic tail. In the lumen of the gut they form into spherical structures called
micelles with a hydrophilic outer layer and hydrophobic centre.
Micelles then become attached to the brush border of enterocytes (intestinal epithelial cells) and they fuse with the cell membrane, allowing fatty acids and glycerol, etc., to enter the cell by diffusion.
The contents of the micelles then become enclosed in a vesicle, and re-form into triglycerides.
These vesicles join onto the Golgi body, and the triglycerides are processed. Triglycerides are combined with protein from the endoplasmic reticulum to form lipoprotein. They are packaged into spherical structures called
chylomicrons, which are taken to the basal surface of the cell and released by exocytosis into the interstitial space inside the central section of the villus.
From here the chylomicrons enter the lymphatic capillary (lacteal) to be taken into the rest of the lymphatic system.
A micelle shown in cross section
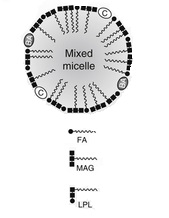
Micelles are about 5 nm in diameter.
They have an outer hydrophilic layer embedded with bile salts (BA) and cholesterol (C) on the surface. The -OH groups of fatty acids (FA), lysophospholipids (LPL) from phospholipid digestion and monoglyceride - monoacyl glycerol - (MAG), are also aligned with the surface and in contact with the watery fluid which surrounds it in the lumen of the ileum.
The hydrophobic tails of fatty acids and MAG project into the centre of the micelles.
A chylomicron - schematic image
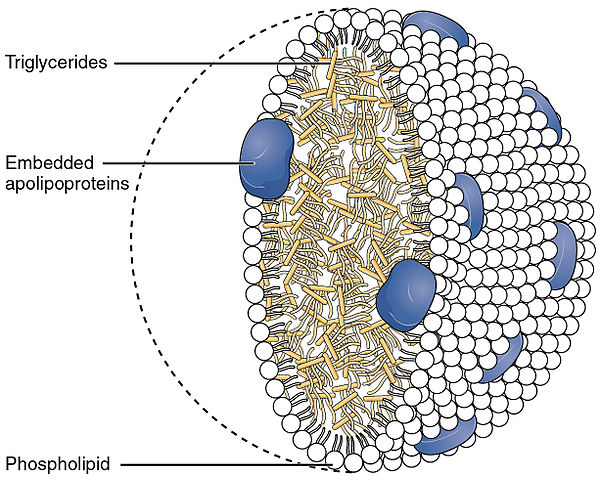 Illustration from Anatomy & Physiology, Connexions Web site. http://cnx.org/content/col11496/1.6/, Jun 19, 2013.
Illustration from Anatomy & Physiology, Connexions Web site. http://cnx.org/content/col11496/1.6/, Jun 19, 2013.
Chylomicrons are also known as ultra low-density lipoproteins (ULDLs).
They vary greatly in size - from 75 to 600 nm in diameter.
They consist of an outer spherical layer of phospholipids, embedded with apolipoproteins, enclosing triglycerides (85-92%), and cholesterol (1-3%). They transport dietary lipids from the intestine to other locations in the body - adipose, cardiac, and skeletal muscle tissue.
Here triglycerides are hydrolysed and free fatty acids are absorbed by the tissues.
Other related topics on this site
(also accessible from the drop-down menu above)
Similar level
Exchanges with the environment
Surface area to volume ratio
Gas exchange
Mass transport in animals
Biological Chemicals
Monomers and polymers - Condensation reactions build up simple molecules into more complex ones
Carbohydrates - Monosaccharides, disaccharides, polysaccharides
Lipids - Fatty acids, triglycerides, phospholipids
Proteins - Amino acids, polypeptides
Enzymes - Hydrolysis and synthesis of biological chemicals all take place under enzyme control
Cells and cellularity
Transport across cell membranes - Digestion products need special mechanisms to enter cells
Response (A2 topic)
Control of blood glucose concentration
Other topics
The human digestive system - Drag n drop revision exercise - not much use if you are using a tablet or mobile
Digestion of starch - practical schedule for an early 6th form practical
Simpler level
How enzymes work - Links to several animations
Class experiments - Quite popular for project work worldwide
The action of amylase on starch - also with 'food tests' on substrates and products
The effect of various factors on enzyme action - more about amylase
Action of pepsin on proteins - using egg white - see the effect of acid
Action of lipase on fats - using milk - see the effect of bile salts
Interactive 3-D molecular graphic models on this site
(also accessible from the drop-down menu above)
The 3-dimensional structure of:
The enzyme amylase
Amylose
Amylopectin
Maltose
Web references
Amylase
Polysaccharides
Structural Basis for Substrate Selectivity in Human Maltase-Glucoamylase and Sucrase-Isomaltase N-terminal Domains*
Trypsin
From Wikipedia, the free encyclopedia

 Home
Home