Prokaryotes are Bacteria,
Cyanobacteria and Archaea.
Bacteria and Cyanobacteria belong to the Domain Prokaryota, whereas Archaea have a Domain of their own.
These are all single-celled organisms, although the cells may remain together after division, producing filaments or aggregations like bunches of grapes. Prokaryotic cells are even smaller than individual cells of eukaryotes.
In fact prokaryotes are usually a similar size to mitochondria of eukaryotes.
Their cells lack a true nucleus, and do not have any membrane-bound organelles.
Bacteria are well known as causative agents for diseases. However there is considerable variation in their metabolism and they can use a number of substrates as a source of energy. As such they perform a number of chemical transformations which are important from an environmental and agricultural perspective, and directly in food production.
Click to see/ hide some examples:
Nitrification - production of nitrate NO3-
(nitrifying bacteria in soil (2 stages):
ammonia NH3 and ammonium NH4+ to nitrite NO2-, Nitrosomonas spp,
nitrite (NO2-) to nitrate Nitrobacter spp)
Nitrogen fixation (free-living bacteria in soil, symbiotic bacteria in root nodules of leguminous plants: Rhizobium leguminosarum)
Denitrification - conversion of nitrate ions into nitrogen gas (anaerobic bacteria in waterlogged soil: Pseudomonas spp and Clostridium spp )
Bacteria in cheese production Lactobacillus species
yoghurt production Lactobacillus + Streptococcus bulgaricus
Acetobacter in production of vinegar
Click to hide the above.
Cyanobacteria have been called blue-green 'algae' because of the blue pigment phycocyanin which they contain in addition to the green chlorophyll a, the same as most green plants, but they are not closely related to algae. Sometimes they are reddish, due to the red pigment phycoerythrin. Some species have gas vesicles for buoyancy, and some are capable of slow gliding movement.
Cyanobacteria are sometimes seen as 'algal blooms' in sea and lakes in warm summer weather, especially if build-up of nitrates and phosphates occurs, possibly resulting from leaching of agricultural fertilisers, or pollution incidents. They produce cyanotoxins which act as
neurotoxins,
hepatotoxins,
cytotoxins, and
endotoxins,
and can be accumulated in shellfish.
Cyanobacteria also carry out useful chemical reactions such as nitrogen fixation as well as photosynthesis.
The importance of cyanobacteria should not be under-estimated in the development of life on this planet over the course of geological time.
Cyanobacteria were the first organisms to carry out photosynthesis about 2.5 billion years (or more) ago. The oxygen they produced was initially chemically absorbed but then it gradually built up in the atmosphere, resulting in the
Great Oxygenation Event.
This changed the balance in microbial life - switching from anaerobic to aerobic dominance - and eventually leading to
the equilibrium between oxygen producers and consumers which is the basis for the ecological dependence of animals on plants today.
The atmospheric oxygen also provided the ozone layer which acted as a protective filter against solar ultra-violet radiation.
Archaea, although less well known generally, also perform various chemical conversions e.g. production of methane. They operate in extreme environmental conditions e.g. volcanic hotsprings, salt lakes.
Three main groups are the halophiles (salt lovers), thermophiles (heat lovers), and acidophiles (acid lovers).
Click on pictures for enlarged version (opens in separate window)
BACTERIA (EUBACTERIA) - false colour scanning electron micrographs
Gram negative Bacterium
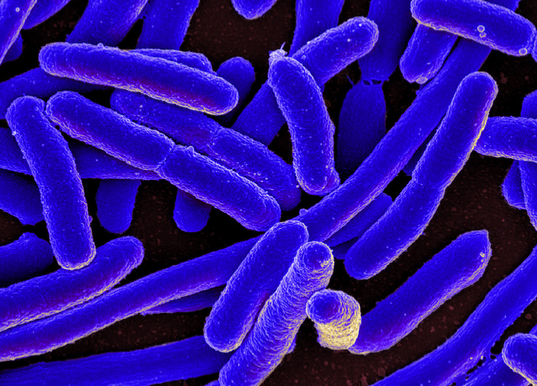 Escherichia coli (E.coli)
Escherichia coli (E.coli)
Size 2 x 0.5 µm
Bacterial cells with a cylindrical shape (rods) are called bacilli.
Spherical cells are called cocci
Gram positive Bacterium
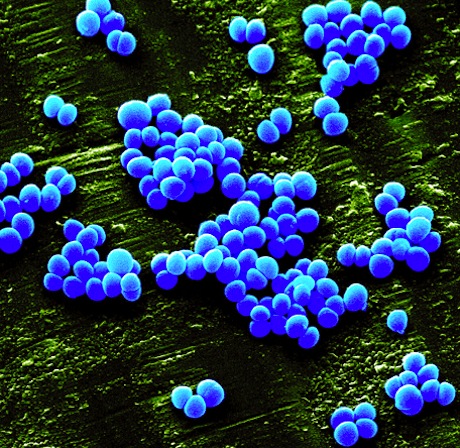 Staphylococcus aureus (MRSA)
Staphylococcus aureus (MRSA)
Diameter 0.6 µm
CYANOBACTERIA (light microscope)
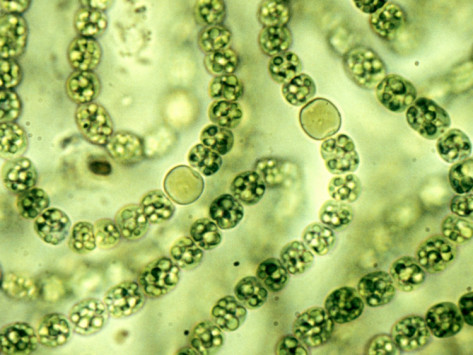 Nostoc
Nostoc
The circular cells visible here are heterocysts, which perform nitrogen fixation in anaerobic conditions.
ARCHAEA (false colour sem)
 Methanosarcina - a methane-producing species
Methanosarcina - a methane-producing species
A section through (part of) a bacterial cell - seen with a transmission electron microscope

Mouseover for labels
Prokaryotic cells contain the following components:
Instead of a nucleus, they have a single
molecule of DNA - actually a circle. This DNA is not associated with proteins to form chromosomes, but it appears as a diffuse mass, free in the cytoplasm.
Prokaryotic cells perform the process of
binary fission in which cells simply divide into two after the DNA has replicated. This does not require special structures like the spindle formed during mitosis in eukaryotes.
Ribosomes are loose in the cytoplasm (not attached to a membrane).
Prokaryotic ribosomes are slightly smaller than eukaryotic ribosomes: approximatly 20 nm in diameter. They are usually described as 70S, with subunits 50S and 30S (Svedberg sedimentation coefficients).
A
cell wall containing
peptidoglycan, also known as
murein, on the outside of a cell membrane.
Peptidoglycan
click to enlarge
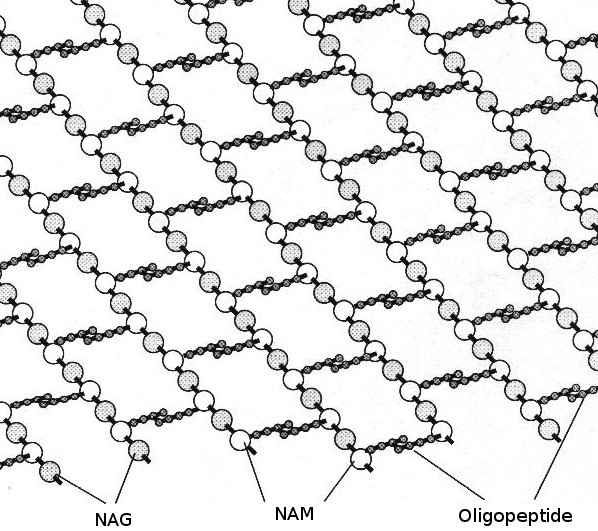
The
glycan structure is rows of substituted glucose-based units:
N-acetyl glucosamine (NAG) alternating with
N-acetyl muramic acid (NAM), linked by β 4 → 1 glycosidic linkages.
Beneath this and extending out sideways from the NAM units are
peptide chains - a few (usually 4 or 5) amino acids linked by peptide linkages (also known as oligopeptides) and cross linked to a similar peptide chain from another glycan row. The actual amino acids vary in different bacterial species and strains - often including unusual "non proteogenic" amino acids, but they usually include both D- and L-alanine.
Gram positive bacteria have several layers of peptidoglycan with lipotechoic acid on the outside - total thickness 20 to 40 nm.
Gram negative bacteria have a single layer of peptidoglycan with an outer lipid membrane on top of that - 2 to 6 nm thick.
This distinction is important as it underlies the different susceptibilities of bacterial types to various antibiotics, e.g. penicillin, and antibacterial enzymes such as lysozyme.
The Gram staining technique emphasises the properties of peptidoglycan: staining with crystal violet (a water-soluble dye) fixing this with Gram's iodine solution, followed by decolorization (using ethanol or acetone), and counterstaining with safranin (a red water-soluble dye).
Gram positive bacteria retain the purple stain whereas Gram negative bacteria lose this and show up as pink.
TS cyanobacterial cell - tem
click to enlarge
") Synechocystis a unicellular cyanobacterium that is very widespread in the marine environment
Synechocystis a unicellular cyanobacterium that is very widespread in the marine environment
Cyanobacteria have cell walls resembling those of gram positive bacteria but with more cross-linking between the peptidoglycan chains, mostly involving the unusual meso-diaminopimelic acid.
On the outside of the peptidoglycan is a layer of proteins fitting together like a mosaic (S-layer) with another layer of protein fibrils, oscillin fibrils, wound helically around the cell.
Inside the cell wall, cyanobacteria have concentric layers of thylakoid membranes which contain photosynthetic pigments rather like chloroplasts in green plants.
These appear to be paired when viewed in section but are probably arranged as flask-shaped structures, one inside another. There is an intermembrane space between these thylakoid membranes.
As well as contributing large amounts of oxygen into the atmosphere from prehistoric time to the present day, cyanobacteria are implicated in the
endosymbiont theory which suggests that chloroplasts of present-day green plants represent an independent organism like them taken into the cell and then incorporated into it.
Archaea do not have true peptidoglycan in their walls. Instead they have pseudopeptidoglycan (also known as pseudomurein). This is composed of N-acetylglucosamine and N-acetyltalosaminuronic acid, linked by
β-1,3-glycosidic bonds.
In addition, their cell membranes have a slightly different molecular architecture based on L-glycerol rather than the R-form which is more normal, and isoprene side chains, often branched, instead of fatty acid chains.
Other components
Some prokaryotes have:
A plasmid
click to enlarge
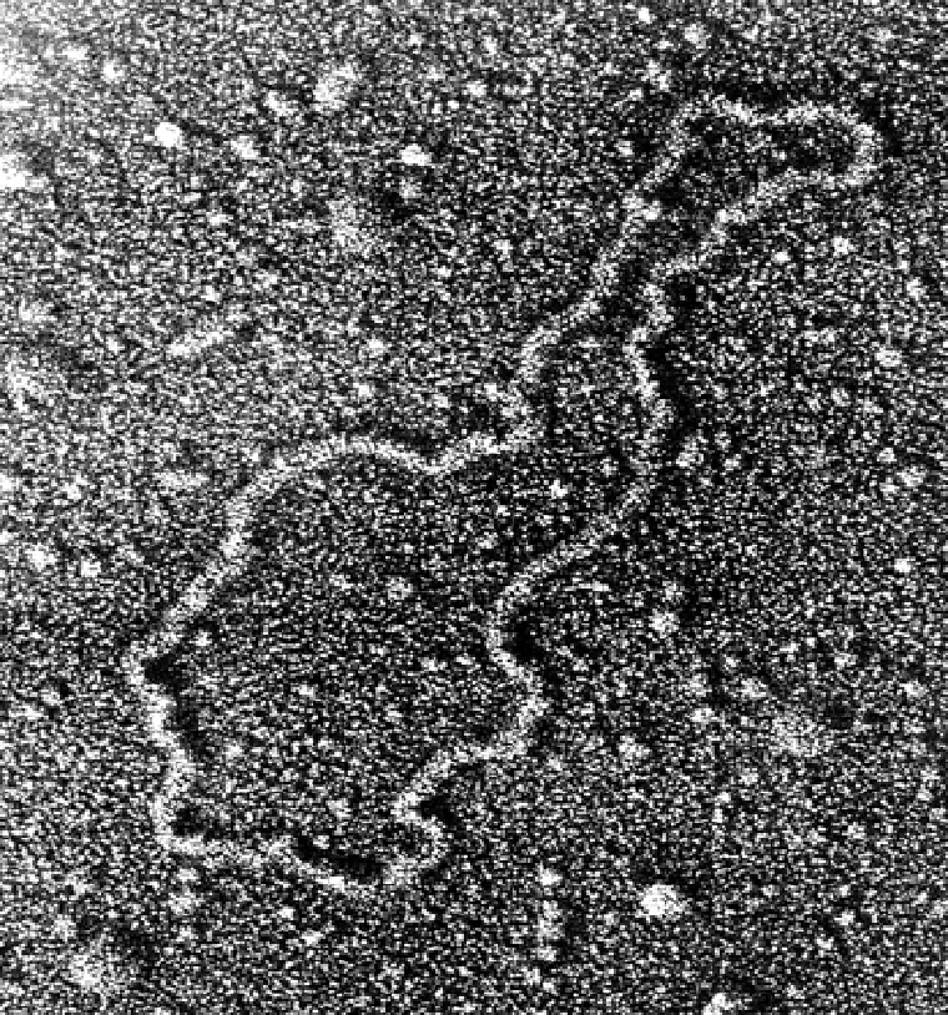 This plasmid has been removed from the bacterial cell and spread onto a flat surface
This plasmid has been removed from the bacterial cell and spread onto a flat surface
- plasmids - extra loops of DNA (distinct from the cell's main DNA) - in the cytoplasm
Genes carried in plasmids may provide bacteria with genetic advantages, such as antibiotic resistance.
When a bacterium divides, all of the plasmids contained within the cell are copied so that each daughter cell receives a copy of each.
Bacteria can also transfer plasmids to one another through a process called conjugation.
Researchers have used plasmids as vectors to clone, transfer, and manipulate genes.
A recombinant plasmid can be produced by inserting DNA fragments or genes into a plasmid.
This plasmid can be introduced into a bacterium by the process called transformation.
Bacteria divide rapidly, and copy DNA fragments in large quantities.
- a capsule surrounding the cell
Encapsulated bacterial cells
click to enlarge
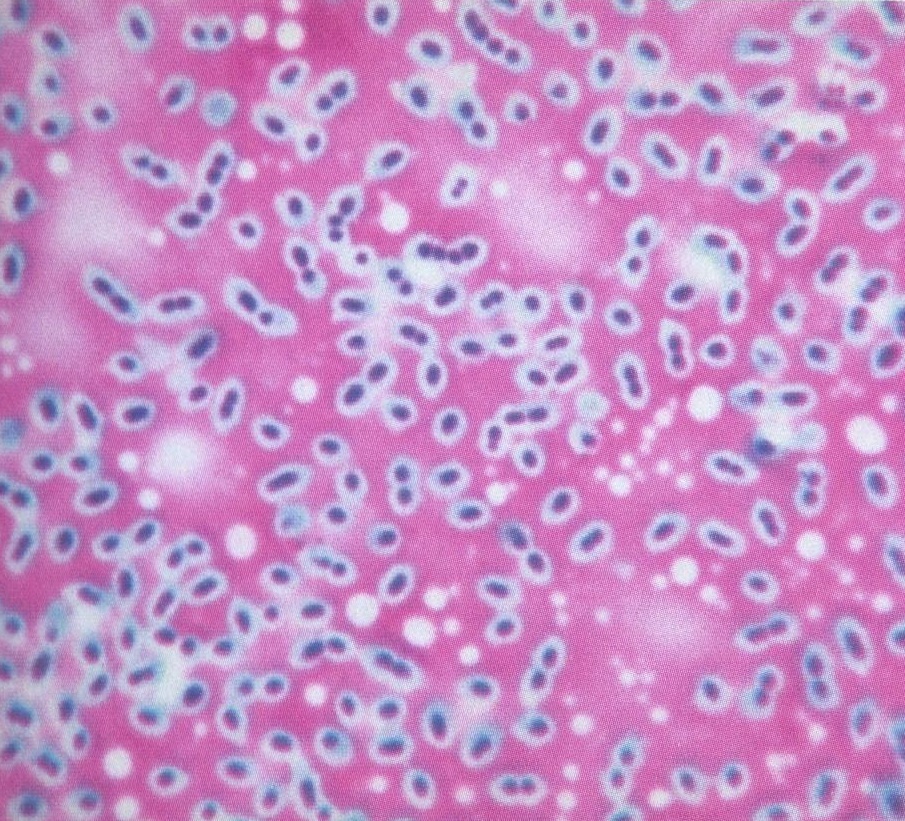 The capsule does not absorb stain so cells have a characteristic halo around them
The capsule does not absorb stain so cells have a characteristic halo around them
This may be composed of a polysaccharide or a polypeptide and it forms a sticky or slimy layer on the outside of the ordinary cell wall and membrane (which is collectively known as the cell envelope).
A capsule may give advantages to a bacterial cell: preventing dehydration by absorbing water and swelling up, or causing it to become attached to surfaces.
Capsules may interfere with viruses (bacteriophages) which attack bacterial cells and can also cover cell-surface antigens which mean that phagocytosis is impaired, so that the body's defence mechanisms are defeated.
As such, the presence of a capsule may be considered to a virulence factor. For example,
there are two forms of Pneumococcus: rough (R) and smooth (S). The S form is more virulent, and produces a capsule. This distinction formed the basis for Frederick Griffith's experiment showing bacterial transformation - an important step in the DNA story, as shown by Oswald Avery, Colin MacLeod, and Maclyn McCarty.
- a flagellum/flagella at one end or all round the cell
A flagellum is a whip-like filament, used as an organ of locomotion.
Bacterial flagella rotate, whereas flagella in eukaryotic cells, e.g. sperm cells, wave from side to side. You could say a prokaryotic flagellum works like a propeller, and a eukaryotic flagellum works like an oar. At the bacterial scale, the viscosity of water is quite important, and forces compound in interesting ways.
 Most diagrams of this structure have been coloured to resemble metalwork!
Most diagrams of this structure have been coloured to resemble metalwork!
Prokaryotic flagella are composed of a protein flagellin. Many subunits of this protein aggregate into a hollow cylindrical structure, which can grow in length by the addition of subunits which move up from below. This is attached to a protein structure called a hook, which passes through ring-shaped proteins like bearings in different outer layers of the cell, and is rotated by a motor-like section embedded in the cell membrane. This is powered by hydrogen ions H+ which build up in the peptidoglycan layer. The motor section is rather similar to the ATPase in mitochondrial and chloroplast membranes, the difference being that ATP is used up in driving round the motor whereas in mitochondria and chloroplasts ATP is accumulated. Some flagella are powered by Na+ gradient.
The rotation of the hook and flagellin filament can be switched to the opposite direction. Surprisingly this does not reverse the direction of movement of the bacterial cell, but causes it to gyrate and tumble in a random fashion. In ideal conditions the bacterial cell moves in a distinctly "forwards" direction, but, if conditions change, the bacterium will change direction until better conditions are regained. Thus the bacterial cell maintains an optimum position in a liquid.




 Home
Home