Action potential
The outer surface of the axon membrane has a positive charge, compared to the inside. When a nervous impulse passes along the axon,
voltage-gated sodium ion channels open, causing
sodium ions to enter by diffusion, down an electrical/chemical gradient. This causes
depolarisation of the membrane
at that point, so that here the inside temporarily has a positive charge - typically about +45 mV.
This means that an electrical circuit exists between this (-ve) region and the neighbouring (+ve) section ("adjoining region") of the axon membrane. This sets up a local current, increasing the permeability of the adjoining region as a result of voltage-gated sodium ion channels opening, allowing sodium ions to enter at that point. In fact the voltage change stimulates more sodium ion channels to open - a
positive feedback effect. At the peak of the action potential, sodium ion channels close. There is a section within the channel called the inactivation gate which plugs the channel, preventing flow of sodium ions. At a later stage when the resting potential is restored, 'deinactivation' occurs and the neurone can transmit another impulse.
Next, voltage-gated potassium ion channels open and potassium ions diffuse out of the axon, down an electrical/chemical gradient, which works in the opposite direction. This has the effect of reversing the axon surface potential back towards its resting (negative) value:
repolarisation.
In fact this reduces the membrane potential below the normal resting potential -
hyperpolarisation - shown as "undershoot".
The potassium ion channels now (gradually) close.
There is a brief period where ions which have recently moved reassort themselves with others on either side of the membrane.
Another action potential cannot pass until the normal resting potential is restored - this gap is called the '
refractory period'. This means that nervous impulses are distinct and discrete pulses, which move in one direction, with distinct gaps (in time) between each.
The depolarisation phase and repolarisation each take about 0.5 ms (1.0 ms total), and the hyperpolarisation phase is often about 3ms, so the maximum frequency is about 250 impulses per second
(250 Hz).

Before and after action potential
This process occurs progressively along the length of the axon, so that the action potential gradually moves along it.
This is called the
propagation of a nervous impulse, because it involves the repeated creation of an identical event, travelling in a set direction.
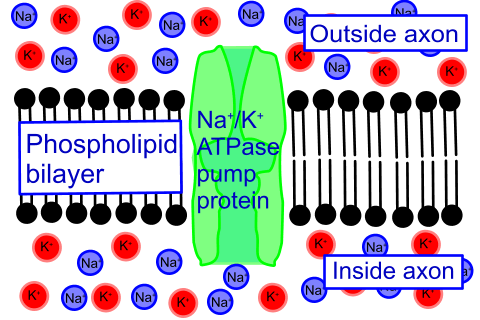
It is sometimes said that after an impulse, the sodium/potassium ion pump can once again pump sodium ions out, and potassium ions in.
However, the
numbers of sodium and potassium ions crossing the membrane at each impulse are insignificant compared with the excess concentrations providing the motive force. It has been shown that (hundreds of) nervous impulses can still be sent after the sodium/potassium ion pump has been inactivated using metabolic inhibitors.
The sodium/potassium ion pump works more or less continuously in the background.
This is the difference it makes
| Ion |
External
concentration |
Internal
concentration |
Equilibrium
potential |
| Na+ |
460 |
50 |
+60mV |
| K+ |
20 |
400 |
-90mV |
| Cl- |
560 |
100 |
-70mV |
| Organic anions |
0 |
370 |
? |
Ionic concentrations (mmol/dm3) on either side of the (squid) axon membrane
and the equilibrium potentials associated with the differences in ionic concentration
The membrane potential can be calculated from the
Goldman-Hodgkin-Katz equation:

where
P
K is the relative membrane permeability for K
+ ions
[K
+]
o is the K
+ ion concentration outside the axon
[K
+]
i is the K
+ ion concentration inside the axon
and similarly for Na
+ and Cl
- ions.
Obviously
R is the universal gas constant (8.314 J.K-1.mol-1)
T is the temperature in Kelvin (K = °C + 273.15)
F is Faraday's constant (96485 C.mol-1)
ln is the log to base e (of the combined fraction)
Since the potassium ion has the highest concentration ratio (inside to outside) and a number of leak channels are open all the time allowing small numbers of potassium ions to leave the axon, it determines the extent of the resting potential - taking it down (about three-quarters of the way) towards the equilibrium potential for potassium. The other ions are negligible at this stage.
During the rising phase of the action potential, permeability to sodium ions rises dramatically as voltage-gated sodium ion channels open, and the membrane potential rises towards the sodium equilibrium potential, reversing the membrane polarity as sodium ions enter the axon.
This is followed by a comparable but extended rise in membrane permeability to potassium ions, again due to voltage-gated ion channels, which allows more potassium ions to move in the opposite direction (out of the axon), taking the membrane potential back down to an even lower value than the resting potential.
Eventually voltage-gated potassium channels close and the membrane is once again dictated by the potassium leak channels.
Chloride ions do not participate in the same way as sodium and potassium ions in nervous impulses along the axon, but they can contribute to inhibitory post-synaptic potentials.
In some neurones and muscle cells, calcium ions contribute to membrane potentials too.















 Home
Home